\documentstyle[etc,html,epsfig,bibtexlogo]{ecal97}
%\documentstyle[bibtexlogo,html,epsfig,twocolumn]{article}
\renewcommand{\topfraction}{1}
\renewcommand{\bottomfraction}{1}
\renewcommand{\textfraction}{0}
\newcommand{\hlnk}[2]{\htmladdnormallink{#1}{#2}}
\newcommand{\hoturl}[1]{\hlnk{\texttt{#1}}{#1}}
\newcommand{\mailto}[1]{\hlnk{\texttt{#1}}{mailto:#1}}
\begin{latexonly}
% Fix hyphenation(s):
\hyphenation{McMullin}
\end{latexonly}
\newcommand{\hole}{\texttt{H}}
\newcommand{\sbst}{\texttt{S}}
\newcommand{\cat}{\texttt{K}}
\newcommand{\lnk}{\texttt{L}}
\newcommand{\lnkplus}{\mbox{\lnk$^{+}$}}
\newcommand{\blnk}{\texttt{BL}}
\begin{document}
\begin{latexonly}
\copyrightdate{1997}
% This must be manually echoed in the .latex2html-init
% also - l2h cannot (perversely) pick it up from here
% in time to put it in the HTML footer...
\end{latexonly}
\newcommand{\dcunumber}{\texttt{bmcm9703}}
\newcommand{\sfinumber}{\texttt{97-02-012}}
\date{February 1997 \\[2mm]
\hlnk{SFI}{http://www.santafe.edu} Working Paper
\sfinumber{}\\
\hlnk{DCU}{http://www.eeng.dcu.ie} Technical Report \dcunumber{}\\[2mm]
\hlnk{\emph{Submitted to ECAL-97, Brighton, UK July
1997.}}{http://www.cogs.susx.ac.uk/ecal97}}
\title{Rediscovering Computational
Autopoiesis}
\author{
\begin{center}
\begin{tabular}[t]{cc}
\makebox[80mm]{\textbf{
\hlnk{Barry McMullin}{http://www.eeng.dcu.ie/\~{}mcmullin}}} &
\makebox[80mm]{\textbf{Francisco J.~Varela}} \\
\hlnk{Santa Fe Institute}{http://www.santafe.edu} &
LENA---CNRS URA 654 \\
1399 Hyde Park Road &
Hopital del la Salpetriere \\
Santa Fe, NM 87501 &
47, Blvd.~de l'Hopital \\
USA &
75651 Paris cedex 13 \\
& France. \\[2mm]
\mailto{mcmullin@eeng.dcu.ie} &
\mailto{fv@ccr.jussieu.fr} \\
\hoturl{http://www.eeng.dcu.ie/\~{}mcmullin} &
\end{tabular}
\end{center}}
\maketitle
%%%% no page number on title page
\thispagestyle{empty}
\vspace*{-7mm}
\begin{abstract}
This paper summarises some initial empirical results
from a new computer model (artificial chemistry)
which exhibits spontaneous emergence and persistence
of \emph{autopoietic} organisation. The model is
based on a system originally presented by Varela,
Maturana and Uribe \cite{Varela:Autopoiesis}. In
carrying out this re-implementation it was found that
an additional interaction (chain-based bond
inhibition), not documented in the original
description by Varela et~al., is critical to the
realisation of the autopoietic phenomena. This
required interaction was re-discovered only following
careful examination of (unpublished) source code for
an early version of the original model. The purpose
of the paper is thus twofold: firstly to identify and
discuss this previously undocumented, but essential,
interaction; and secondly to argue, on the basis of
this particular case, for the importance of
exploiting the emerging technologies which support
publication of completely detailed software models
(in addition, of course, to conventional publication
of summary experimental results).
\end{abstract}
\vfill
\begin{quote}
\textbf{Keywords:} Autopoiesis, Artificial Life,
Artificial Chemistry, Origin of Life.
\end{quote}
\newpage
\section{Introduction}
The original computer model of a minimal autopoietic
system was first described by Varela, Maturana and
Uribe in 1974 \cite{Varela:Autopoiesis}. This
presented experimental data from the model showing both
the spontaneous formation and ongoing repair of an
autopoietic system embedded in a two dimensional,
discrete space. This was accompanied by both a
qualitative description of the artificial chemistry
realised by the model, and a more detailed algorithmic
account of their simulation program.
A number of technical difficulties with interpreting
that original algorithm, and apparent discrepancies
between the algorithm and the experimental data, have
been discussed in a previously published working paper
\cite{McMullin:OriginalAutop}. That paper also
incorporates, as an appendix, the \texttt{FORTRAN-IV}
code of a version of the original program used by
Varela et~al. Careful study of this code has allowed
the identification of an additional interaction that
was not explicitly identified in the original
description of the model.
In this paper we present experimental results from a
completely new implementation of the qualitative
chemistry described by Varela et~al.\fsp which suggests
that this additional interaction is, indeed, critical
to the realisation of the autopoietic phenomena; and
that, conversely, provided this additional interaction
is included, the autopoietic phenomena are not
dependent on any particular details of the original
program or algorithm, but may be expected in \emph{any}
system sharing the same qualitative chemistry.
\section{The Original Qualitative Chemistry\label{original}}
The chemistry takes places in a discrete, two
dimensional, space. Each position in the space is
either empty or occupied by a single particle.
Particles generally move in random walks in the space.
There are three distinct particle types, engaging in
three distinct reactions:
\begin{itemize}
\item Production: Two substrate (\sbst) particles may react, in
the presence of a catalyst (\cat) particle to form a
link (\lnk) particle.\par
\item Bonding: \lnk{} particles may bond to other
\lnk{} particles. Each \lnk{} particle can form (at
most) two bonds, thus allowing the formation of
indefinitely long chains, which may close to form
membranes. Bonded \lnk{} particles become
immobile.\par
\item Disintegration: An \lnk{} particle may
spontaneously disintegrate, yielding two \sbst{}
particles. When this occurs any bonds associated with
the \lnk{} particle are destroyed also.
\end{itemize}
Chains of \lnk{} particles are permeable to \sbst{}
particles but impermeable to \cat{} and \lnk{}
particles. Thus a closed chain, or membrane, which
encloses \cat{} or \lnk{} particles effectively traps
such particles.
\section{The Phenomena}
The basic autopoietic phenomenon predicted for this
system is the possibility of realising dynamic
cell-like structures which, on an ongoing basis,
produce the conditions for their own maintenance. Such
a system would consist of a closed chain (membrane) of \lnk{}
particles enclosing one or more \cat{} particles.
Because \sbst{} particles can permeate through the
membrane, there can be ongoing production of \lnk{}
particles. Since these cannot escape from the
membrane, this will result in the build up of a
relatively high concentration of \lnk{} particles. On
an ongoing basis, the membrane will rupture as a result
of disintegration of component \lnk{} particles.
Because of the high concentration of \lnk{} particles
inside the membrane, there should be a high probability
that one of these will drift to the rupture site and
effect a repair, \emph{before} the \cat{} particle(s)
escape, thus re-establishing precisely the conditions
allowing the build up of that high concentration of
\lnk{} particles.
A secondary phenomenon which \emph{may} arise is the
spontaneous establishment of an autopoietic system from
a randomised initial arrangement of the particles.
Clearly, the issue of spontaneous formation does not
arise unless the system actually supports
autopoietic organisation. In this sense the phenomenon
of autopoietic organisation
is \emph{logically} prior to spontaneous formation (though
chronologically following from it). For this reason,
the phenomenon of spontaneous formation will \emph{not}
be considered further in this paper. Instead, in all
the experiments reported, a putatively autopoietic
entity will be artificially introduced into the system;
the question at issue will be whether this entity
succeeds in realising the autopoietic reaction network
already described.
\section{The SCL Program}
The newly developed program is called SCL (for
Substrate-Catalyst-Link) \cite{McMullin:SCL}. This has
been implemented using the
SWARM\footnote{\hoturl{http://www.santafe.edu/projects/swarm}}
simulation system, developed at the Santa Fe
Institute\footnote{\hoturl{http://www.santafe.edu}}.
A conscious decision was taken that SCL would
\emph{not} be based on the \emph{algorithm} originally
published by Varela et~al., but should rather reflect
an independent implementation of the same
\emph{qualitative} chemistry. This stemmed partly from
the previously documented problems with the original
algorithm \cite{McMullin:OriginalAutop}; but it also
reflected a desire to test the robustness of the
autopoietic phenomena---i.e.\fsp are they perhaps reliant
on some artifact of the original program and/or
algorithm, or are they robust outcomes from the given
qualitative chemistry.
The version of SCL used to generate the results
described in this paper (v0.05) differs in minor ways
from that described in the previously published
documentation (v0.04) \cite{McMullin:SCL}. The
complete source code relating to SCL v0.05 is
available as:
\begin{quote}
\varurl
{ftp://ftp.santafe.edu/pub/swarm/
\indent users-contrib/anarchy/scl-0.05.tar.gz}
{ftp://ftp.santafe.edu/pub/swarm/users-contrib/anarchy/scl-0.05.tar.gz}
\end{quote}
The SCL data files relating to the experiments
described here are available in:
\begin{quote}
\varurl
{ftp://ftp.santafe.edu/pub/swarm/
\indent users-contrib/anarchy/scl-data00.tar.gz}
{ftp://ftp.santafe.edu/pub/swarm/users-contrib/anarchy/scl-data00.tar.gz}
\end{quote}
A key to the display of the three particle types in SCL
is provided in figure~\ref{key}.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{key.ps}}
\end{latexonly}
\end{center}
\caption{Key to Particle Types.}
\label{key}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
\section{Experimental Protocol}
This paper will describe the results from two separate
experiments with SCL. In each case the same
experimental protocol was followed. Each experiment
consisted of 5 runs of SCL. The initial configuration
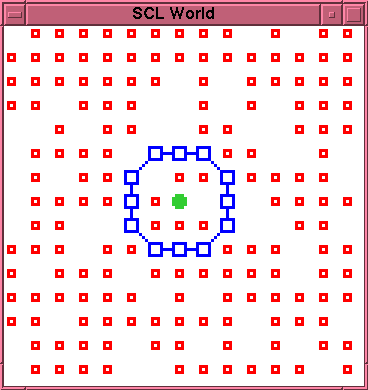
was identical in all runs, and is illustrated in
figure~\ref{initial}. This comprises a single
artificially constructed cell-like entity, being a
closed membrane of \lnk{} particles enclosing a single
\cat{} particle. This is embedded in a $15 \times 15$
toroidal space. The five runs in each experiment
differed only in the initial state of the underlying
pseudo-random number generator. These five distinct
initial states are specified in the files
\texttt{run1.stt} through \texttt{run5.stt} in the
distributed data file archive
(\texttt{scl-data00.tar.gz}).
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{initial.ps}}
\end{latexonly}
\end{center}
\caption{Initial Configuration.}
\label{initial}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
\section{Experimental Protocol}
This paper will describe the results from two separate
experiments with SCL. In each case the same
experimental protocol was followed. Each experiment
consisted of 5 runs of SCL. The initial configuration
was identical in all runs, and is illustrated in
figure~\ref{initial}. This comprises a single
artificially constructed cell-like entity, being a
closed membrane of \lnk{} particles enclosing a single
\cat{} particle. This is embedded in a $15 \times 15$
toroidal space. The five runs in each experiment
differed only in the initial state of the underlying
pseudo-random number generator. These five distinct
initial states are specified in the files
\texttt{run1.stt} through \texttt{run5.stt} in the
distributed data file archive
(\texttt{scl-data00.tar.gz}).
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{initial.ps}}
\end{latexonly}
\end{center}
\caption{Initial Configuration.}
\label{initial}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
The two experiments differed only in that experiment~1
implemented just the reactions of the qualitative
chemistry described in section~\ref{original}, whereas
experiment~2 incorporated the newly rediscovered
chain-based bond inhibition interaction (to be
discussed in section~\ref{inhibition}).
SCL supports a variety of parameters controlling
reaction rates, mobility parameters, etc. Apart from
the parameter controlling the additional interaction
just mentioned, these parameters were held constant
across all runs and both experiments. These parameter
sets for experiments~1 and~2 are specified in the files
\texttt{exp1.prm} and \texttt{exp2.prm} respectively,
in the distributed data file archive.
The \texttt{disintegrationProbability} parameter was
set at 0.001 in all cases.\footnote{This is a factor of 10
smaller than the value originally suggested by Varela
et~al.\fsp This reflects the fact that the
\emph{maximum} rate of the production reaction is
approximately this much slower in SCL than in the
original model, so that 10 timesteps in SCL can be
considered roughly comparable to one timestep in the
original.} This is the probability that any given
\lnk{} particle will disintegrate per unit time. The
membrane in the initial configuration is composed of 12
\lnk{} particles. It follows that the expected time to
first rupture of the initial membrane is given by:
%\begin{eqnarray*}
%\tau & = & \frac{1}{1 - (1 - P_d)^{12}} \\
%& \simeq & 84
%\end{eqnarray*}
\[ \tau = \frac{1}{1 - (1 - P_d)^{12}} \simeq 84 \]
\section{Experiment 1}
\subsection{Run 1-1}
As expected, \sbst{} particles initially permeate
through the membrane and, under the influence of the
\cat{} particle, production of \lnk{} particles starts.
However, instead of these \lnk{} particles remaining
mobile, trapped within the membrane, in readiness to
repair any rupture, they begin to spontaneously bond to
\emph{each other}. Given that bonded \lnk{} particles
are specified to be immobile, this means that such
particles are \emph{not} available to drift to a
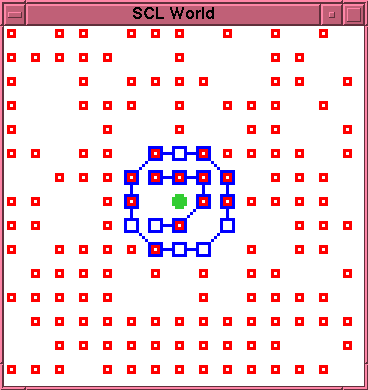
rupture site. The screenshot of
figure~\ref{run1-1-0110} was taken at time 110. The
membrane has not yet suffered any decay. However, the
interior of the membrane is now completely clogged with
bonded---and thus immobile---\lnk{} particles. Only two
open positions remain inside the membrane, one occupied
by the \cat{} particle. Since the production reaction
requires two \sbst{} particles adjacent to each other
and to the \cat{} particle, there is no longer any
available site for further production within the
membrane, and further production of \lnk{} particles is
impossible. It follows that, whenever the membrane
does eventually rupture, there will be no mobile \lnk{}
particles available to effect a repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-1-0110.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 1, Time 110.}
\label{run1-1-0110}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
The two experiments differed only in that experiment~1
implemented just the reactions of the qualitative
chemistry described in section~\ref{original}, whereas
experiment~2 incorporated the newly rediscovered
chain-based bond inhibition interaction (to be
discussed in section~\ref{inhibition}).
SCL supports a variety of parameters controlling
reaction rates, mobility parameters, etc. Apart from
the parameter controlling the additional interaction
just mentioned, these parameters were held constant
across all runs and both experiments. These parameter
sets for experiments~1 and~2 are specified in the files
\texttt{exp1.prm} and \texttt{exp2.prm} respectively,
in the distributed data file archive.
The \texttt{disintegrationProbability} parameter was
set at 0.001 in all cases.\footnote{This is a factor of 10
smaller than the value originally suggested by Varela
et~al.\fsp This reflects the fact that the
\emph{maximum} rate of the production reaction is
approximately this much slower in SCL than in the
original model, so that 10 timesteps in SCL can be
considered roughly comparable to one timestep in the
original.} This is the probability that any given
\lnk{} particle will disintegrate per unit time. The
membrane in the initial configuration is composed of 12
\lnk{} particles. It follows that the expected time to
first rupture of the initial membrane is given by:
%\begin{eqnarray*}
%\tau & = & \frac{1}{1 - (1 - P_d)^{12}} \\
%& \simeq & 84
%\end{eqnarray*}
\[ \tau = \frac{1}{1 - (1 - P_d)^{12}} \simeq 84 \]
\section{Experiment 1}
\subsection{Run 1-1}
As expected, \sbst{} particles initially permeate
through the membrane and, under the influence of the
\cat{} particle, production of \lnk{} particles starts.
However, instead of these \lnk{} particles remaining
mobile, trapped within the membrane, in readiness to
repair any rupture, they begin to spontaneously bond to
\emph{each other}. Given that bonded \lnk{} particles
are specified to be immobile, this means that such
particles are \emph{not} available to drift to a
rupture site. The screenshot of
figure~\ref{run1-1-0110} was taken at time 110. The
membrane has not yet suffered any decay. However, the
interior of the membrane is now completely clogged with
bonded---and thus immobile---\lnk{} particles. Only two
open positions remain inside the membrane, one occupied
by the \cat{} particle. Since the production reaction
requires two \sbst{} particles adjacent to each other
and to the \cat{} particle, there is no longer any
available site for further production within the
membrane, and further production of \lnk{} particles is
impossible. It follows that, whenever the membrane
does eventually rupture, there will be no mobile \lnk{}
particles available to effect a repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-1-0110.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 1, Time 110.}
\label{run1-1-0110}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
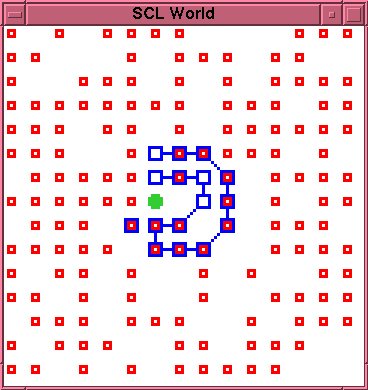
In fact, the membrane suffers a double rupture at times 234 and
235, yielding the configuration shown in figure~\ref{run1-1-0235}.
The chain which had previously been formed inside the
membrane now becomes spliced to one side of the rupture site,
forming a folded chain. This no longer encloses the \cat{}
particle. Indeed, should the folded chain become closed,
the \cat{} particle would necessarily be \emph{outside} it.
Thus, the initial, putatively autopoietic, entity has clearly
now irreversibly degenerated, without having undergone even a
single episode of self-repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-1-0235.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 1, Time 235.}
\label{run1-1-0235}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
In fact, the membrane suffers a double rupture at times 234 and
235, yielding the configuration shown in figure~\ref{run1-1-0235}.
The chain which had previously been formed inside the
membrane now becomes spliced to one side of the rupture site,
forming a folded chain. This no longer encloses the \cat{}
particle. Indeed, should the folded chain become closed,
the \cat{} particle would necessarily be \emph{outside} it.
Thus, the initial, putatively autopoietic, entity has clearly
now irreversibly degenerated, without having undergone even a
single episode of self-repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-1-0235.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 1, Time 235.}
\label{run1-1-0235}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
\subsection{Run 1-2}
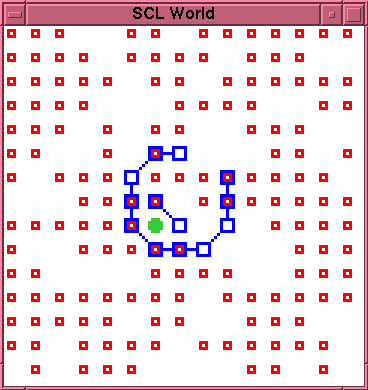
On this run, the initial rupture of the membrane occurs
relatively early, at time 31. Just one \lnk{} particle has
been produced within the membrane by this time. However,
since this means the particle is still mobile, there is
at least some possibility that it may drift to the
rupture site and effect a repair. A second \lnk{} particle
is produced at time 63, thus improving the possibility
of a repair. However, at time 69, these two \lnk{}
particles bond to each other, thus becoming immobile,
and unavailable to drift to the rupture site. Moreover,
they are located in such a position that the \cat{}
particle is blocked in all four directions. This
configuration is illustrated in figure~\ref{run1-2-0069}.
Not only is the \cat{} particle now also effectively
immobile, but, again, there is no space available
adjacent to it to permit further production of new
\lnk{} particles. Thus, there is no possibility of
repairing the existing rupture. As with run~1,
the initial entity has clearly
already irreversibly degenerated.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-2-0069.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 2, Time 069.}
\label{run1-2-0069}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
\subsection{Run 1-2}
On this run, the initial rupture of the membrane occurs
relatively early, at time 31. Just one \lnk{} particle has
been produced within the membrane by this time. However,
since this means the particle is still mobile, there is
at least some possibility that it may drift to the
rupture site and effect a repair. A second \lnk{} particle
is produced at time 63, thus improving the possibility
of a repair. However, at time 69, these two \lnk{}
particles bond to each other, thus becoming immobile,
and unavailable to drift to the rupture site. Moreover,
they are located in such a position that the \cat{}
particle is blocked in all four directions. This
configuration is illustrated in figure~\ref{run1-2-0069}.
Not only is the \cat{} particle now also effectively
immobile, but, again, there is no space available
adjacent to it to permit further production of new
\lnk{} particles. Thus, there is no possibility of
repairing the existing rupture. As with run~1,
the initial entity has clearly
already irreversibly degenerated.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-2-0069.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 2, Time 069.}
\label{run1-2-0069}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
\subsection{Runs 1-3 to 1-5}
Given the descriptions of runs~1 and~2, only a brief description of
the remaining runs is necessary. Precisely the same failure
mechanism is again observed: the \lnk{} particles produced
within the membrane spontaneously bond to each other, thus
becoming immobile, rather than remaining available to drift
to a rupture site when it arises; the interior of the membrane
becomes progressively clogged up, until there is no longer
space available for further production. At this point,
since no \lnk{} particles are available to repair any rupture,
and no more can be produced within the membrane, the original
entity has effectively degenerated. In all three runs this occurs
without even a single episode of successful repair of the
membrane. The times at which this condition is reached are as follows:
\begin{center}
\begin{tabular}{ll}
Run & Time \\
3 & 282 \\
4 & 126 \\
5 & 165
\end{tabular}
\end{center}
These terminal configurations for runs~3--5 are shown
in figures~\ref{run1-3-0282},
\ref{run1-4-0126}~and~\ref{run1-5-0165} respectively.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-3-0282.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 3, Time 282.}
\label{run1-3-0282}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
\subsection{Runs 1-3 to 1-5}
Given the descriptions of runs~1 and~2, only a brief description of
the remaining runs is necessary. Precisely the same failure
mechanism is again observed: the \lnk{} particles produced
within the membrane spontaneously bond to each other, thus
becoming immobile, rather than remaining available to drift
to a rupture site when it arises; the interior of the membrane
becomes progressively clogged up, until there is no longer
space available for further production. At this point,
since no \lnk{} particles are available to repair any rupture,
and no more can be produced within the membrane, the original
entity has effectively degenerated. In all three runs this occurs
without even a single episode of successful repair of the
membrane. The times at which this condition is reached are as follows:
\begin{center}
\begin{tabular}{ll}
Run & Time \\
3 & 282 \\
4 & 126 \\
5 & 165
\end{tabular}
\end{center}
These terminal configurations for runs~3--5 are shown
in figures~\ref{run1-3-0282},
\ref{run1-4-0126}~and~\ref{run1-5-0165} respectively.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-3-0282.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 3, Time 282.}
\label{run1-3-0282}
\end{figure}
\begin{rawhtml}
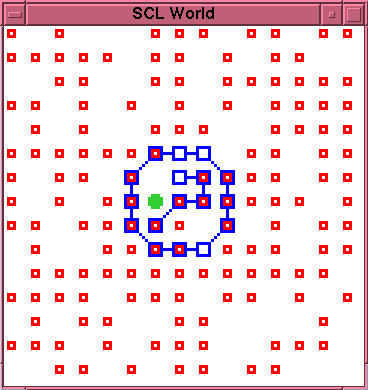 \end{rawhtml}
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-4-0126.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 4, Time 126.}
\label{run1-4-0126}
%\vspace*{30mm}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-4-0126.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 4, Time 126.}
\label{run1-4-0126}
%\vspace*{30mm}
\end{figure}
\begin{rawhtml}
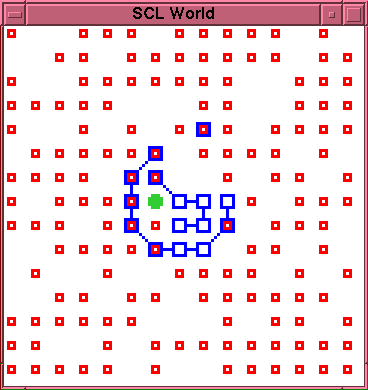 \end{rawhtml}
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-5-0165.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 5, Time 165.}
\label{run1-5-0165}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run1-5-0165.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 1, Run 5, Time 165.}
\label{run1-5-0165}
\end{figure}
\begin{rawhtml}
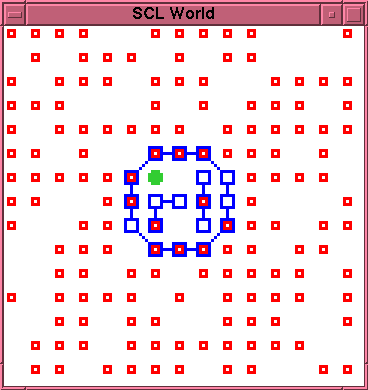 \end{rawhtml}
\subsection{Discussion 1}
In all five runs of experiment~1 a consistent
\emph{failure} of the autopoietic process was observed.
This is due to the spontaneous and premature bonding of
the \lnk{} particles produced within the membrane, thus
making them immobile and unavailable to effect a repair
to the membrane. With the benefit of these
experimental results, it seems fairly obvious that this
failure mode was already implicit in the
\emph{qualitative} chemistry described by Varela
et~al.\fsp \cite{Varela:Autopoiesis}. It is evidently
\emph{not} dependent on any particular details of the
implementation, nor on the specific parameters
settings.
This conclusion is corroborated by the fact that this
same failure mechanism has been observed in previous
(unpublished) experiments with two other,
\emph{independent}, implementations of this reaction
scheme \cite{jullien:SAP,mingers:SAP}, and has also
been previously reported by Lizana
\cite{Lizana:Autopoiesis}. This class of failure seems
also to have been recognised, at least implicitly, in
the re-implementation(s) carried out by Milan Zeleny
\cite[Figure~4]{Zeleny:SelfOrganization}.
Two attempted solutions to this failure mechanism
were briefly investigated, before the preferred
solution, to be discussed in section~\ref{inhibition}, was
finally identified.
Firstly, the bonding reaction was separated into two
cases: bonding between two free \lnk{} particles, and
bonding between a free \lnk{} particle and an \lnk{}
particle already having one bond. The latter is the
case of interest for membrane repair. These were
controlled by separate rate parameters. This allowed
the ``spontaneous'' bonding reaction to be made very
slow. This should ensure that the free \lnk{}
particles formed within the membrane would not
spontaneously bond with each other but would rather be
held in reserve for membrane repair.\footnote{Of
course, this would make spontaneous formation of an
initial membrane much less probable; but that issue
was deferred.} However, this idea proved largely
ineffective. The problem is that, once a rupture
\emph{does} occur it frequently happens that, instead
of a single free \lnk{} repairing the membrane,
\emph{all} of the free \lnk{} particles become quickly
incorporated into an inward spiraling chain fragment.
The second mechanism was separately (?) suggested by
both Zeleny \cite{Zeleny:SelfOrganization} and Lizana
\cite{Lizana:Autopoiesis}. This involves inhibiting
bonding to a free \lnk{} particle in some neighborhood
of any \cat{} particle. In Lizana's case, this effect
seems to have been limited to the \emph{immediate}
(Moore) neighborhood of a \cat{} particle, whereas
Zeleny seems to have used arbitrarily large (and
dynamically changing?) neighborhoods. The idea appears
to be that the \cat{} particle(s) can establish zone(s)
of bond inhibition around them. The membrane can then
form (roughly) at the edge of such a zone. \lnk{}
particles within these zones will remain free, and
ready to drift to a rupture site to effect a repair.
Both Zeleney and Lizana apparently got this mechanism
to give somewhat satisfactory results. The mechanism
has been investigated to only a limited extent with
SCL. Specifically, the use of indefinitely large
inhibition zones (as suggested by Zeleny) has \emph{not}
been pursued, since it violates an objective that the
model should rely only on local (Moore neighborhood)
interactions. With this (self-imposed) restriction, the
results have generally been mediocre. Two
counteracting effects have been noticed. Firstly, even
within a relatively small membrane such as illustrated
in figure~\ref{initial}, the \cat{} particle may
transiently drift away from the central position; if
free \lnk{} particles also drift away from this
position, then they may still be able to spontaneously
bond and become immobile. Even though the \cat{}
particle may now drift back into their vicinity, it is
now too late---the bonding has already
occurred.\footnote{Presumably, the \cat{} particle
should not be assigned an effect of rupturing these bonds
again, because they cannot be distinguished from the
bonds making up the membrane; on the other hand, this
does seem to have been a mechanism actually used by
Zeleny in some experiments
\cite[Figure~6]{Zeleny:SelfOrganization}.}
Secondly, if a rupture occurs in the neighborhood of
the \cat{} particle it is now very difficult to effect
a repair, even if a free \lnk{} particle should drift
into an appropriate position; worse still, this is
precisely the situation in which swift repair is most
important, lest the \cat{} particle should escape
completely. These problems can be overcome, to an
extent, by making the \cat{} particle immobile (in the
center of the cavity). While Lizana's description is
not fully detailed, it seems that this may be what she
indeed did. The mobility of \cat{} particles also seems
to have been severely constrained in a number of
Zeleny's experiments. In our view, this significantly
reduces the generality and interest of the model, and
must be considered an unsatisfactory solution.
\section{Chain-based Bond Inhibition\label{inhibition}}
By far the most troubling aspect of the results
discussed above is that they are not
consistent with the experimental results originally
presented by Varela et~al.\fsp
\cite{Varela:Autopoiesis}.
In particular, a careful examination of those original
results suggests that the model must have had some,
unspecified, mechanism to overcome or preempt the class
of failure now described here. However, given that the
work was done over 25 years ago, it seemed that it
would be extremely difficult to gain much further
insight into this problem. The current author who was
involved in the original work (Varela), no longer had
any clear recollection of what additional mechanism was
present in the model to account for this discrepancy.
Fortunately, a printout of an early version of the
original simulation program, coded in FORTRAN-IV, has
recently been rediscovered, and has now been
incorporated in a published technical report
\cite{McMullin:OriginalAutop}. As discussed in more
detail in that report, this program has been rekeyed,
and it has been possible to execute it again. This did
not reproduce the precise results of the original
publication; most likely it was not exactly the same
version, and, in any case, the original random number
source is no longer available, so the precise execution
trajectory is bound to be different. However, this did
suffice to show that the program did, indeed, exhibit
some mechanism whereby free links, confined within the
membrane, tended \emph{not} to spontaneously bond to
each other. This motivated a detailed reanalysis of
the program code, which finally resulted in the
identification of a previously un-reported
interaction---\emph{chain-based bond inhibition}.
This is an interaction whereby bonding is inhibited to
any free \lnk{} particle which is in the immediate
vicinity of another \lnk{} particle which is doubly
bonded. In effect then, a free \lnk{} particle cannot
form a bond as long as it is alongside (as opposed to
at the end of) an existing chain of \lnk{} particles;
but it \emph{can} form bond(s) when it is at the end of
a chain; and, especially, when it is positioned at a
site where a chain has broken (i.e.\fsp a rupture
site).\footnote{This interaction has previously been
outlined in the SCL documentation
\cite{McMullin:SCL}. However, there is an error, or
ambiguity, in that description, in that it suggests
that bond inhibition applies to both free and singly
bonded \lnk{} particles. In fact, it applies only to
free \lnk{} particles. Applying it to singly bonded
\lnk{} particles would actually \emph{prevent}
membrane repair from taking place.}
The next section reviews experimental results from SCL
when the chain-based bond inhibition reaction is enabled.
\section{Experiment 2}
\subsection{Run 2-1}
Between time 0 and time 226 the initial membrane
suffers two ruptures which are repaired with no change
of membrane morphology. Between time 227 and 444 there
are four rupture and repair episodes, yielding the new
membrane morphology shown in
figure~\ref{run2-1-0444}.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-1-0444.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 1, Time 444.}
\label{run2-1-0444}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
\subsection{Discussion 1}
In all five runs of experiment~1 a consistent
\emph{failure} of the autopoietic process was observed.
This is due to the spontaneous and premature bonding of
the \lnk{} particles produced within the membrane, thus
making them immobile and unavailable to effect a repair
to the membrane. With the benefit of these
experimental results, it seems fairly obvious that this
failure mode was already implicit in the
\emph{qualitative} chemistry described by Varela
et~al.\fsp \cite{Varela:Autopoiesis}. It is evidently
\emph{not} dependent on any particular details of the
implementation, nor on the specific parameters
settings.
This conclusion is corroborated by the fact that this
same failure mechanism has been observed in previous
(unpublished) experiments with two other,
\emph{independent}, implementations of this reaction
scheme \cite{jullien:SAP,mingers:SAP}, and has also
been previously reported by Lizana
\cite{Lizana:Autopoiesis}. This class of failure seems
also to have been recognised, at least implicitly, in
the re-implementation(s) carried out by Milan Zeleny
\cite[Figure~4]{Zeleny:SelfOrganization}.
Two attempted solutions to this failure mechanism
were briefly investigated, before the preferred
solution, to be discussed in section~\ref{inhibition}, was
finally identified.
Firstly, the bonding reaction was separated into two
cases: bonding between two free \lnk{} particles, and
bonding between a free \lnk{} particle and an \lnk{}
particle already having one bond. The latter is the
case of interest for membrane repair. These were
controlled by separate rate parameters. This allowed
the ``spontaneous'' bonding reaction to be made very
slow. This should ensure that the free \lnk{}
particles formed within the membrane would not
spontaneously bond with each other but would rather be
held in reserve for membrane repair.\footnote{Of
course, this would make spontaneous formation of an
initial membrane much less probable; but that issue
was deferred.} However, this idea proved largely
ineffective. The problem is that, once a rupture
\emph{does} occur it frequently happens that, instead
of a single free \lnk{} repairing the membrane,
\emph{all} of the free \lnk{} particles become quickly
incorporated into an inward spiraling chain fragment.
The second mechanism was separately (?) suggested by
both Zeleny \cite{Zeleny:SelfOrganization} and Lizana
\cite{Lizana:Autopoiesis}. This involves inhibiting
bonding to a free \lnk{} particle in some neighborhood
of any \cat{} particle. In Lizana's case, this effect
seems to have been limited to the \emph{immediate}
(Moore) neighborhood of a \cat{} particle, whereas
Zeleny seems to have used arbitrarily large (and
dynamically changing?) neighborhoods. The idea appears
to be that the \cat{} particle(s) can establish zone(s)
of bond inhibition around them. The membrane can then
form (roughly) at the edge of such a zone. \lnk{}
particles within these zones will remain free, and
ready to drift to a rupture site to effect a repair.
Both Zeleney and Lizana apparently got this mechanism
to give somewhat satisfactory results. The mechanism
has been investigated to only a limited extent with
SCL. Specifically, the use of indefinitely large
inhibition zones (as suggested by Zeleny) has \emph{not}
been pursued, since it violates an objective that the
model should rely only on local (Moore neighborhood)
interactions. With this (self-imposed) restriction, the
results have generally been mediocre. Two
counteracting effects have been noticed. Firstly, even
within a relatively small membrane such as illustrated
in figure~\ref{initial}, the \cat{} particle may
transiently drift away from the central position; if
free \lnk{} particles also drift away from this
position, then they may still be able to spontaneously
bond and become immobile. Even though the \cat{}
particle may now drift back into their vicinity, it is
now too late---the bonding has already
occurred.\footnote{Presumably, the \cat{} particle
should not be assigned an effect of rupturing these bonds
again, because they cannot be distinguished from the
bonds making up the membrane; on the other hand, this
does seem to have been a mechanism actually used by
Zeleny in some experiments
\cite[Figure~6]{Zeleny:SelfOrganization}.}
Secondly, if a rupture occurs in the neighborhood of
the \cat{} particle it is now very difficult to effect
a repair, even if a free \lnk{} particle should drift
into an appropriate position; worse still, this is
precisely the situation in which swift repair is most
important, lest the \cat{} particle should escape
completely. These problems can be overcome, to an
extent, by making the \cat{} particle immobile (in the
center of the cavity). While Lizana's description is
not fully detailed, it seems that this may be what she
indeed did. The mobility of \cat{} particles also seems
to have been severely constrained in a number of
Zeleny's experiments. In our view, this significantly
reduces the generality and interest of the model, and
must be considered an unsatisfactory solution.
\section{Chain-based Bond Inhibition\label{inhibition}}
By far the most troubling aspect of the results
discussed above is that they are not
consistent with the experimental results originally
presented by Varela et~al.\fsp
\cite{Varela:Autopoiesis}.
In particular, a careful examination of those original
results suggests that the model must have had some,
unspecified, mechanism to overcome or preempt the class
of failure now described here. However, given that the
work was done over 25 years ago, it seemed that it
would be extremely difficult to gain much further
insight into this problem. The current author who was
involved in the original work (Varela), no longer had
any clear recollection of what additional mechanism was
present in the model to account for this discrepancy.
Fortunately, a printout of an early version of the
original simulation program, coded in FORTRAN-IV, has
recently been rediscovered, and has now been
incorporated in a published technical report
\cite{McMullin:OriginalAutop}. As discussed in more
detail in that report, this program has been rekeyed,
and it has been possible to execute it again. This did
not reproduce the precise results of the original
publication; most likely it was not exactly the same
version, and, in any case, the original random number
source is no longer available, so the precise execution
trajectory is bound to be different. However, this did
suffice to show that the program did, indeed, exhibit
some mechanism whereby free links, confined within the
membrane, tended \emph{not} to spontaneously bond to
each other. This motivated a detailed reanalysis of
the program code, which finally resulted in the
identification of a previously un-reported
interaction---\emph{chain-based bond inhibition}.
This is an interaction whereby bonding is inhibited to
any free \lnk{} particle which is in the immediate
vicinity of another \lnk{} particle which is doubly
bonded. In effect then, a free \lnk{} particle cannot
form a bond as long as it is alongside (as opposed to
at the end of) an existing chain of \lnk{} particles;
but it \emph{can} form bond(s) when it is at the end of
a chain; and, especially, when it is positioned at a
site where a chain has broken (i.e.\fsp a rupture
site).\footnote{This interaction has previously been
outlined in the SCL documentation
\cite{McMullin:SCL}. However, there is an error, or
ambiguity, in that description, in that it suggests
that bond inhibition applies to both free and singly
bonded \lnk{} particles. In fact, it applies only to
free \lnk{} particles. Applying it to singly bonded
\lnk{} particles would actually \emph{prevent}
membrane repair from taking place.}
The next section reviews experimental results from SCL
when the chain-based bond inhibition reaction is enabled.
\section{Experiment 2}
\subsection{Run 2-1}
Between time 0 and time 226 the initial membrane
suffers two ruptures which are repaired with no change
of membrane morphology. Between time 227 and 444 there
are four rupture and repair episodes, yielding the new
membrane morphology shown in
figure~\ref{run2-1-0444}.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-1-0444.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 1, Time 444.}
\label{run2-1-0444}
\end{figure}
\begin{rawhtml}
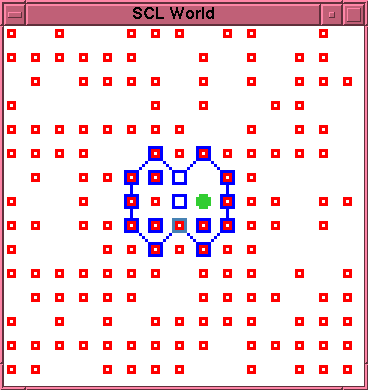 \end{rawhtml}
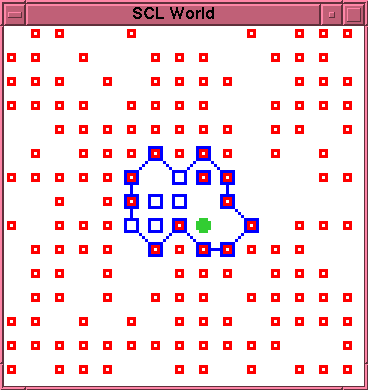
This new morphology appears to be relatively robust.
The entity persists in this morphology up to time 1250,
in the course of which there are 5 more rupture and
(successful) repair episodes. Between time 1250 and
time 1310 there are two rupture and repair episodes
yielding the new membrane morphology shown in
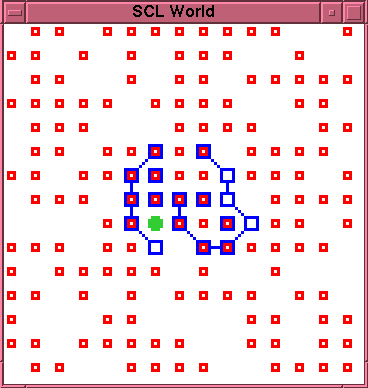
figure~\ref{run2-1-1310}. The entity survives in this
morphology, through two more rupture and repair
episodes until time 1741. There are then two ruptures
in quick succession, at times 1742 and 1745. At time
1746 the membrane fragments, and partially spirals into
the cavity, as shown in figure~\ref{run2-1-1746}. It
is then no longer possible to recover the closed
membrane through any simple process of self repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-1-1310.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 1, Time 1310.}
\label{run2-1-1310}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
This new morphology appears to be relatively robust.
The entity persists in this morphology up to time 1250,
in the course of which there are 5 more rupture and
(successful) repair episodes. Between time 1250 and
time 1310 there are two rupture and repair episodes
yielding the new membrane morphology shown in
figure~\ref{run2-1-1310}. The entity survives in this
morphology, through two more rupture and repair
episodes until time 1741. There are then two ruptures
in quick succession, at times 1742 and 1745. At time
1746 the membrane fragments, and partially spirals into
the cavity, as shown in figure~\ref{run2-1-1746}. It
is then no longer possible to recover the closed
membrane through any simple process of self repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-1-1310.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 1, Time 1310.}
\label{run2-1-1310}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-1-1746.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 1, Time 1746.}
\label{run2-1-1746}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-1-1746.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 1, Time 1746.}
\label{run2-1-1746}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
\bigskip
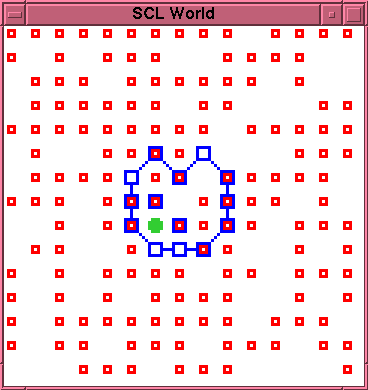
\subsection{Run 2-2}
Between time 0 and time 133 the initial membrane
suffers three ruptures which are repaired with no
change of membrane morphology. A rupture at time 134 is
repaired at time 137, yielding the new membrane
morphology shown in figure~\ref{run2-2-0137}. The
entity persists in this morphology up to time 452, in
the course of which there is one more rupture and
(successful) repair episode. A further rupture at time
453 is eventually repaired at time 555; but in the
interim, a second rupture at time 542 leads to a
partial spiral into the cavity, as with run~1, and
again it is then no longer possible to recover a closed
membrane through any simple process of self repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-2-0137.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 2, Time 137.}
\label{run2-2-0137}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
\bigskip
\subsection{Run 2-2}
Between time 0 and time 133 the initial membrane
suffers three ruptures which are repaired with no
change of membrane morphology. A rupture at time 134 is
repaired at time 137, yielding the new membrane
morphology shown in figure~\ref{run2-2-0137}. The
entity persists in this morphology up to time 452, in
the course of which there is one more rupture and
(successful) repair episode. A further rupture at time
453 is eventually repaired at time 555; but in the
interim, a second rupture at time 542 leads to a
partial spiral into the cavity, as with run~1, and
again it is then no longer possible to recover a closed
membrane through any simple process of self repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-2-0137.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 2, Time 137.}
\label{run2-2-0137}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
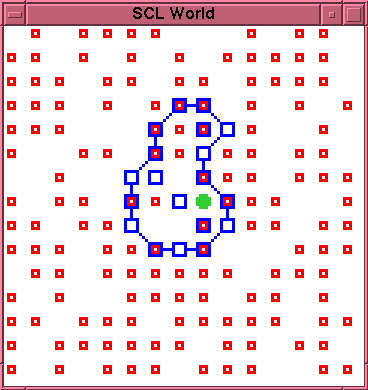
\subsection{Run 2-3}
In this run there are two very early ruptures (times 6
and 13), before there has been time for an effective
build up in the concentration of free \lnk{} particles.
The \cat{} particle almost escapes immediately, but,
instead, an extension of the membrane forms around it.
There is a further independent rupture at time 139, but
at time 148 a closed membrane reforms with the new
morphology shown in figure~\ref{run2-3-0148}
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-3-0148.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 3, Time 148.}
\label{run2-3-0148}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
\subsection{Run 2-3}
In this run there are two very early ruptures (times 6
and 13), before there has been time for an effective
build up in the concentration of free \lnk{} particles.
The \cat{} particle almost escapes immediately, but,
instead, an extension of the membrane forms around it.
There is a further independent rupture at time 139, but
at time 148 a closed membrane reforms with the new
morphology shown in figure~\ref{run2-3-0148}
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-3-0148.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 3, Time 148.}
\label{run2-3-0148}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
A further rupture at time 171 results again in an
inward spiral and it is then no longer possible to
recover a closed membrane through any simple process of
self repair.
\subsection{Run 2-4}
Following three rupture and repair episodes, at time
199 the entity forms into the same morphology
encountered in run~1
(figure~\ref{run2-1-0444})---albeit, now rotated
through $90^\circ$. This morphology again appears
relatively robust, persisting from time 199 to time
1437, through 12 rupture and self-repair episodes.
Between times 1438 and 1500 there are \emph{four}
additional ruptures. Of these, one is successfully
repaired, but the overall damage to the membrane is now
too great, and by time 1508 it has degenerated into a
single curved chain as shown in
figure~\ref{run2-4-1508}. Again, it is then no longer
possible to recover a closed membrane through any
simple process of self repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-4-1508.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 4, Time 1508.}
\label{run2-4-1508}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
A further rupture at time 171 results again in an
inward spiral and it is then no longer possible to
recover a closed membrane through any simple process of
self repair.
\subsection{Run 2-4}
Following three rupture and repair episodes, at time
199 the entity forms into the same morphology
encountered in run~1
(figure~\ref{run2-1-0444})---albeit, now rotated
through $90^\circ$. This morphology again appears
relatively robust, persisting from time 199 to time
1437, through 12 rupture and self-repair episodes.
Between times 1438 and 1500 there are \emph{four}
additional ruptures. Of these, one is successfully
repaired, but the overall damage to the membrane is now
too great, and by time 1508 it has degenerated into a
single curved chain as shown in
figure~\ref{run2-4-1508}. Again, it is then no longer
possible to recover a closed membrane through any
simple process of self repair.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-4-1508.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 4, Time 1508.}
\label{run2-4-1508}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
\subsection{Run 2-5}
In this run there are two early ruptures at time 22 and
119, severely damaging the original membrane. An inward
spiral forms. Coincidentally, another rupture allows
the inward spiral to close forming a ``new'' membrane
at time 245, with the morphology shown in
figure~\ref{run2-5-0245}. However, the cavity is now
linear and thus does not afford \emph{any} reaction
sites for production of new \lnk{} particles. This
entity is therefore not capable of re-establishing the
autopoietic reaction network.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-5-0245.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 5, Time 245.}
\label{run2-5-0245}
\end{figure}
\begin{rawhtml}
\end{rawhtml}
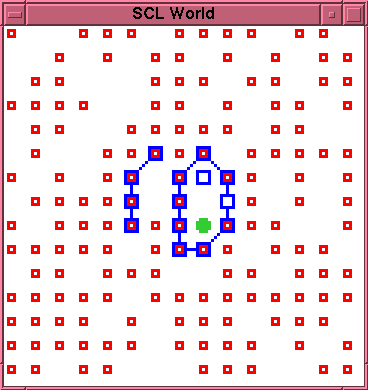
\subsection{Run 2-5}
In this run there are two early ruptures at time 22 and
119, severely damaging the original membrane. An inward
spiral forms. Coincidentally, another rupture allows
the inward spiral to close forming a ``new'' membrane
at time 245, with the morphology shown in
figure~\ref{run2-5-0245}. However, the cavity is now
linear and thus does not afford \emph{any} reaction
sites for production of new \lnk{} particles. This
entity is therefore not capable of re-establishing the
autopoietic reaction network.
\begin{figure}[tb]
\begin{center}
\begin{latexonly}
\mbox{\epsffile{run2-5-0245.ps}}
\end{latexonly}
\end{center}
\caption{Experiment 2, Run 5, Time 245.}
\label{run2-5-0245}
\end{figure}
\begin{rawhtml}
 \end{rawhtml}
\subsection{Discussion 2}
There is substantial variation among the five runs
comprising experiment~2. In runs~3 and~5, the initial
entity effectively fails completely to establish a
closed, autopoietic, reaction network. However, in
runs~1, 2~and~4, an autopoietic reaction network
\emph{is} established, and a succession of successful
repair episodes occurs. In runs 1~and~4 a morphology
becomes established which is apparently particularly
robust, persisting in each case for approximately 1000
time steps of the model.
The work reported here has not involved any extensive
or comprehensive investigation of variations in the
various reaction rate and mobility parameters available
in the SCL model. It might well be possible to find
combinations of parameter settings in which the
establishment and maintenance of autopoietic reaction
networks is more robust, and the autopoietic entities
would thus be more stable and longer lived. However,
the basic results of experiment~2 clearly show that
this model \emph{can} exhibit persistent, self
repairing, autopoietic reaction networks, in the form
originally described by Varela et~al.\fsp
\cite{Varela:Autopoiesis}
\begin{quote}
\emph{Given that the only difference between
experiment~1 and experiment~2 is the (re-)introduction
of the chain-based bond inhibition interaction, it
seems reasonable to conclude that this phenomenon of
computational autopoiesis relies critically on the
presence of this interaction.}
\end{quote}
\section{Conclusion}
The primary conclusion from the work described here is
that the original report of computational modelling of
autopoiesis \cite{Varela:Autopoiesis} was flawed, in
that it failed to identify the chain-based bond
inhibition interaction as being present and, indeed, as
being an essential requirement for the achievement of
the described autopoietic phenomena. Given the lapse
of time since the original publication, it is now
difficult to suggest any definitive explanation as to
how this interaction, actually present in the program
code, came to be overlooked in the qualitative and
algorithmic descriptions. However, as described
elsewhere \cite{Varela:EarlyDays}, the work was carried
out during a difficult and turbulent time in Chile,
and, further, there was a considerable time interval
between the actual experiments and eventual
publication. These factors together probably provide
an adequate explanation for the oversight.
In any case, the work described here also raises a more
general question about the publication of
computationally based ALife research. A key feature of
scientific publication is that it should facilitate
independent critical testing of whatever phenomena are
presented. In this particular case, the defect in the
original reporting (\emph{not} a defect in the original
model!) was uncovered only when a copy of the original
program code was rediscovered by chance. At the time
of the original publication, the technological
facilities were not generally available to support easy
distribution or access to accompanying code---but this
is no longer the case. We would suggest therefore that
as a general principle, published reports on computer
models of ALife should be accompanied by access to the
program code for the models on the World Wide Web.
Bare access to program code is, of course, of limited
value in itself. Effective critical review would
require that it should be ``reasonably'' feasible that
others in the community be able to \emph{execute} (and,
indeed, modify) this code. This suggests a need for
some degree of standardisation, where that is possible.
The Swarm simulation system, with its open licensing
for scientific research, offers a candidate platform
for such standardisation. Indeed, this was a key
reason for adopting Swarm in the development of the SCL
system \cite{McMullin:SCL}. Our experience of using
Swarm in this application suggests that it can provide
a stable, efficient, and portable basis for wide
dissemination of this kind of ALife research.
\section*{Acknowledgments\label{acknowledge}}
Initial work on re-implementing a computational model
of autopoiesis was carried out by a former student of
McMullin, Hyder Aswad. We am also grateful to
Francoise Jullien and John Mingers for making
available their independent re-implementations of the
model of \cite{Varela:Autopoiesis}. Ms.~Marita Prandoni
and Ms.~Peggy Jones provided invaluable assistance in
translating original documents from Spanish.
All errors in the current paper remain,
of course, our own responsibility.
This paper was written while McMullin was a visiting
researcher (1996/97) with the \hlnk{Swarm Project
Group}{http://www.santafe.edu/projects/swarm} at the
\hlnk{Santa Fe Institute}{http://www.santafe.edu}.
This visit was made possible by generous support from
\hlnk{Dublin City University}{http://www.dcu.ie} and
the Swarm Project. Swarm has been supported by Grant
No.~N00014-94-1-G014 from the Department of the Navy,
Naval Research Laboratory, acting in cooperation with
the Advanced Research Projects Agency. Swarm has also
benefited from earlier support from The Carol O'Donnell
Foundation and from Mr.~and Mrs.~Michael Grantham.
%\newpage
\section*{Revision History\label{revision}}
First published in February 1997, simultaneously as
Working Paper No.~\sfinumber{} of the \hlnk{Santa Fe
Institute}{http://www.santafe.edu} and as Technical
Report No.~\dcunumber{} of the \hlnk{School of
Electronic Engineering}{http://www.eeng.dcu.ie} of
\hlnk{Dublin City University}{http://www.dcu.ie}.
Submitted to \hlnk{ECAL-97, Brighton, UK July
1997}{http://www.cogs.susx.ac.uk/ecal97}.
\nocite{Maturana:AutopoiesisAndCognition}
\bibliographystyle{ecal97}
\bibliography{bmcm9703}
\section*{Retrieval\label{retrieval}}
The resources comprising this paper are electronically
retrievable, in various formats, via the \hlnk{World Wide Web}
{http://www.w3.org/hypertext/WWW/TheProject}, from
either of the following URL's:
\begin{center}
\noindent
\texttt{\varurl{http://www.santafe.edu/sfi/publications/
Working-Papers/97-02-012/}
{http://www.santafe.edu/sfi/publications/Working-Papers/97-02-012/}}\\
\noindent\hoturl{http://www.eeng.dcu.ie/\~{}autonomy/bmcm9703/}
\end{center}
Either of these URL's will directly access the root
node of the HTML format paper. Alternatively, an
additional filename may be appended, to retrieve one of
the other formats, or related resources, as follows:
\begin{itemize}
\item The complete set of files making up the HTML format of the
paper is available in the single compressed archive file
\hoturl{bmcm9703.html.tar.gz} (c.~102~KByte),
which can be conveniently downloaded for local and/or offline
browsing.
\item A printable version of the paper is available in
postscript format as file \hoturl{bmcm9703.ps}
(c.~637~KByte, 10~pages) or in compressed form as
file \hoturl{bmcm9703.ps.gz} (c.~96~KByte).
\item The original \LaTeX{} source of the paper is
available as file \hoturl{bmcm9703.tex}
(c.~40~KByte) or in compressed form as file
\hoturl{bmcm9703.tex.gz} (c.~13~KByte).
\item The \BibTeX{} source for the reference
list is available as file \hoturl{bmcm9703.bib}
(c.~6~KByte).
\item A complete set of all the files just listed
is available as a single compressed
archive file \hoturl{bmcm9703.tar.gz}
(c.~209~KByte).
\item The source code distribution of SCL is available
in the single compressed archive file
\hoturl{scl-0.05.tar.gz} (c.~34~KByte).
\item The data file archive referred to in the paper is available
in the single compressed archive file
\hoturl{scl-data00.tar.gz} (c.~1~KByte).
\end{itemize}
\section*{Copyright\label{copyright}}
This paper is copyright \copyright \thecopyrightdate{}
by Barry McMullin and Francisco J.~Varela.
Permission is hereby granted to private individuals to
access, copy and distribute this work, for purposes of
private study only, provided that the distribution is
complete and unmodified, is accompanied by this
copyright notice, and that no charges are levied;
multiple copying for class use in educational
organisations is specifically \emph{permitted}.
The work may \emph{not\/} be accessed or copied, in
whole or in part, for commercial purposes, except with
the prior written permission of the authors.
All other rights reserved.
\end{document}
\end{rawhtml}
\subsection{Discussion 2}
There is substantial variation among the five runs
comprising experiment~2. In runs~3 and~5, the initial
entity effectively fails completely to establish a
closed, autopoietic, reaction network. However, in
runs~1, 2~and~4, an autopoietic reaction network
\emph{is} established, and a succession of successful
repair episodes occurs. In runs 1~and~4 a morphology
becomes established which is apparently particularly
robust, persisting in each case for approximately 1000
time steps of the model.
The work reported here has not involved any extensive
or comprehensive investigation of variations in the
various reaction rate and mobility parameters available
in the SCL model. It might well be possible to find
combinations of parameter settings in which the
establishment and maintenance of autopoietic reaction
networks is more robust, and the autopoietic entities
would thus be more stable and longer lived. However,
the basic results of experiment~2 clearly show that
this model \emph{can} exhibit persistent, self
repairing, autopoietic reaction networks, in the form
originally described by Varela et~al.\fsp
\cite{Varela:Autopoiesis}
\begin{quote}
\emph{Given that the only difference between
experiment~1 and experiment~2 is the (re-)introduction
of the chain-based bond inhibition interaction, it
seems reasonable to conclude that this phenomenon of
computational autopoiesis relies critically on the
presence of this interaction.}
\end{quote}
\section{Conclusion}
The primary conclusion from the work described here is
that the original report of computational modelling of
autopoiesis \cite{Varela:Autopoiesis} was flawed, in
that it failed to identify the chain-based bond
inhibition interaction as being present and, indeed, as
being an essential requirement for the achievement of
the described autopoietic phenomena. Given the lapse
of time since the original publication, it is now
difficult to suggest any definitive explanation as to
how this interaction, actually present in the program
code, came to be overlooked in the qualitative and
algorithmic descriptions. However, as described
elsewhere \cite{Varela:EarlyDays}, the work was carried
out during a difficult and turbulent time in Chile,
and, further, there was a considerable time interval
between the actual experiments and eventual
publication. These factors together probably provide
an adequate explanation for the oversight.
In any case, the work described here also raises a more
general question about the publication of
computationally based ALife research. A key feature of
scientific publication is that it should facilitate
independent critical testing of whatever phenomena are
presented. In this particular case, the defect in the
original reporting (\emph{not} a defect in the original
model!) was uncovered only when a copy of the original
program code was rediscovered by chance. At the time
of the original publication, the technological
facilities were not generally available to support easy
distribution or access to accompanying code---but this
is no longer the case. We would suggest therefore that
as a general principle, published reports on computer
models of ALife should be accompanied by access to the
program code for the models on the World Wide Web.
Bare access to program code is, of course, of limited
value in itself. Effective critical review would
require that it should be ``reasonably'' feasible that
others in the community be able to \emph{execute} (and,
indeed, modify) this code. This suggests a need for
some degree of standardisation, where that is possible.
The Swarm simulation system, with its open licensing
for scientific research, offers a candidate platform
for such standardisation. Indeed, this was a key
reason for adopting Swarm in the development of the SCL
system \cite{McMullin:SCL}. Our experience of using
Swarm in this application suggests that it can provide
a stable, efficient, and portable basis for wide
dissemination of this kind of ALife research.
\section*{Acknowledgments\label{acknowledge}}
Initial work on re-implementing a computational model
of autopoiesis was carried out by a former student of
McMullin, Hyder Aswad. We am also grateful to
Francoise Jullien and John Mingers for making
available their independent re-implementations of the
model of \cite{Varela:Autopoiesis}. Ms.~Marita Prandoni
and Ms.~Peggy Jones provided invaluable assistance in
translating original documents from Spanish.
All errors in the current paper remain,
of course, our own responsibility.
This paper was written while McMullin was a visiting
researcher (1996/97) with the \hlnk{Swarm Project
Group}{http://www.santafe.edu/projects/swarm} at the
\hlnk{Santa Fe Institute}{http://www.santafe.edu}.
This visit was made possible by generous support from
\hlnk{Dublin City University}{http://www.dcu.ie} and
the Swarm Project. Swarm has been supported by Grant
No.~N00014-94-1-G014 from the Department of the Navy,
Naval Research Laboratory, acting in cooperation with
the Advanced Research Projects Agency. Swarm has also
benefited from earlier support from The Carol O'Donnell
Foundation and from Mr.~and Mrs.~Michael Grantham.
%\newpage
\section*{Revision History\label{revision}}
First published in February 1997, simultaneously as
Working Paper No.~\sfinumber{} of the \hlnk{Santa Fe
Institute}{http://www.santafe.edu} and as Technical
Report No.~\dcunumber{} of the \hlnk{School of
Electronic Engineering}{http://www.eeng.dcu.ie} of
\hlnk{Dublin City University}{http://www.dcu.ie}.
Submitted to \hlnk{ECAL-97, Brighton, UK July
1997}{http://www.cogs.susx.ac.uk/ecal97}.
\nocite{Maturana:AutopoiesisAndCognition}
\bibliographystyle{ecal97}
\bibliography{bmcm9703}
\section*{Retrieval\label{retrieval}}
The resources comprising this paper are electronically
retrievable, in various formats, via the \hlnk{World Wide Web}
{http://www.w3.org/hypertext/WWW/TheProject}, from
either of the following URL's:
\begin{center}
\noindent
\texttt{\varurl{http://www.santafe.edu/sfi/publications/
Working-Papers/97-02-012/}
{http://www.santafe.edu/sfi/publications/Working-Papers/97-02-012/}}\\
\noindent\hoturl{http://www.eeng.dcu.ie/\~{}autonomy/bmcm9703/}
\end{center}
Either of these URL's will directly access the root
node of the HTML format paper. Alternatively, an
additional filename may be appended, to retrieve one of
the other formats, or related resources, as follows:
\begin{itemize}
\item The complete set of files making up the HTML format of the
paper is available in the single compressed archive file
\hoturl{bmcm9703.html.tar.gz} (c.~102~KByte),
which can be conveniently downloaded for local and/or offline
browsing.
\item A printable version of the paper is available in
postscript format as file \hoturl{bmcm9703.ps}
(c.~637~KByte, 10~pages) or in compressed form as
file \hoturl{bmcm9703.ps.gz} (c.~96~KByte).
\item The original \LaTeX{} source of the paper is
available as file \hoturl{bmcm9703.tex}
(c.~40~KByte) or in compressed form as file
\hoturl{bmcm9703.tex.gz} (c.~13~KByte).
\item The \BibTeX{} source for the reference
list is available as file \hoturl{bmcm9703.bib}
(c.~6~KByte).
\item A complete set of all the files just listed
is available as a single compressed
archive file \hoturl{bmcm9703.tar.gz}
(c.~209~KByte).
\item The source code distribution of SCL is available
in the single compressed archive file
\hoturl{scl-0.05.tar.gz} (c.~34~KByte).
\item The data file archive referred to in the paper is available
in the single compressed archive file
\hoturl{scl-data00.tar.gz} (c.~1~KByte).
\end{itemize}
\section*{Copyright\label{copyright}}
This paper is copyright \copyright \thecopyrightdate{}
by Barry McMullin and Francisco J.~Varela.
Permission is hereby granted to private individuals to
access, copy and distribute this work, for purposes of
private study only, provided that the distribution is
complete and unmodified, is accompanied by this
copyright notice, and that no charges are levied;
multiple copying for class use in educational
organisations is specifically \emph{permitted}.
The work may \emph{not\/} be accessed or copied, in
whole or in part, for commercial purposes, except with
the prior written permission of the authors.
All other rights reserved.
\end{document}