The question I now propose to address is this:
What entities, or kinds of entities, can qualify as the "units of selection" in Darwinian evolution?
This issue has received extended consideration, as summarised by, for example, Hull (1981). Not unnaturally, this discussion has been carried out mainly in the particular context of biological evolution, but Hull remarks that the issue is so fundamental that it deserves to be called `metaphysical'. Similarly, Dawkins has suggested that the question is not simply one of empirically deciding which real entities function as units of selection, but rather is a "dispute about about what we ought to mean when we talk about a unit of selection" (Dawkins 1982b).
Needless to say, given the intense scholarly attention which has been devoted to this issue, I do not claim to have a definitive resolution to offer. However, I shall at least try to offer a clear target for further criticism.
In particular, I embrace Dawkins' view that the substantive
question here is to elucidate "the nature of the entity for
whose benefit adaptations may be said to exist"
(Dawkins 1982a, p. 81).![]() This, whatever it is, is the entity which I shall identify as the
"unit of selection" .
This, whatever it is, is the entity which I shall identify as the
"unit of selection" .
To make the question even more precise, I am interested in whether the unit of selection is an actor or a lineage. I shall suggest that, in certain, simple, cases, there may be very little difference between the two; and that this explains why the two possibilities are commonly confused. However, I shall conclude that, in the general case, the two possibilities are very different; and that the correct candidate is, quite unambiguously, a particular kind of lineage.
If "adaptation" evolves at all as a result of Darwinian
processes![]() then it does so in the form of a change (or accumulation of
changes) which, in each case, allows a new lineage to
selectively displace an old one because it has higher fitness. I
take it that there is no substantive dispute about this. Darwinian
selection is clearly not a process of "displacement" of one
actor by another. Similarly, while selection is more usually
described in terms of "populations" displacing each other,
these so-called "populations" are, in fact, not just
any old groupings of actors (no matter how alike), but groupings
which are established by virtue of common ancestry - which is
to say, lineages.
then it does so in the form of a change (or accumulation of
changes) which, in each case, allows a new lineage to
selectively displace an old one because it has higher fitness. I
take it that there is no substantive dispute about this. Darwinian
selection is clearly not a process of "displacement" of one
actor by another. Similarly, while selection is more usually
described in terms of "populations" displacing each other,
these so-called "populations" are, in fact, not just
any old groupings of actors (no matter how alike), but groupings
which are established by virtue of common ancestry - which is
to say, lineages.
By lineage here I mean simply the set of all descendents of a given founder (as already indicated in Fig.). Granted, it is only certain kinds of lineages which can participate in selection; these are lineages which are characterised by some selectively important attribute (or "trait" ). I shall distinguish these by the term Similarity Lineage or S-lineage. Briefly, consider some actor which (through mutation or some other mechanism) is different from its parent(s) in some way which is potentially selectively significant. Then I call that actor an S-founder - founder of an S-lineage. The S-lineage is the set of all (and only) those descendents of the S-founder which are "similar" to the S-founder in respect of this trait or character (whether "expressed" or just as "carriers" ). See Fig..
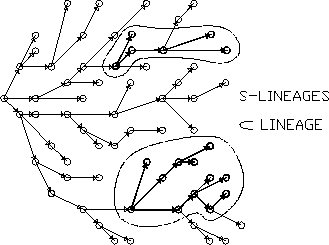
Figure 2: An S-lineage is a subset of a lineage.
The S-lineage concept relates to at least some of the common biological definitions of "species" - but can also potentially map onto both higher and lower taxonomic categories! A fundamental distinction from any conventional "species" concept is that S-lineages can, and generally will, intersect: a particular individual actor may be a member of numerous, distinct, S-lineages - even S-lineages which are competing with each other. Clearly, the concept is a moderately complex one, which I cannot elaborate fully here. A more detailed discussion is available in (McMullin 1992a, Section 5).
In any case, it follows from this that "adaptations" which are the subject of Darwinian selection must be such as to favour an S-lineage relative to its competitors. This immediately suggests the view that adaptations can be viewed as Darwinian only to the extent that they are beneficial to the S-lineage exhibiting them (at least for the duration of the selection episode): i.e. that S-lineages are the "units of selection" . However, this needs to be considered in somewhat more depth.
In the simplest case, fitness can be improved by effects which are uniform and consistent for all members of an S-lineage. This kind of effect is particularly easy to envisage, given that all actors in a given S-lineage are already expected to be "similar" to a greater or lesser extent, simply by virtue of their common membership of that putative S-lineage.
Ultimately, this effect would be manifested in one of just two
possible ways: if the actors uniformly![]() live longer than the actors of the competing S-lineage(s), then the
mortality (expected death rate) of the S-lineage will be less than
its competitors; or if the actors all uniformly have more
offspring (per unit time) then the fecundity (expected birth
rate) of the S-lineage will be greater than its competitors. In
either case, the size of the S-lineage would be expected to
increase relative to its competitor(s); and if the difference
were large enough, and maintained long enough, the S-lineage would
displace its competitors essentially deterministically - which is
to say, an episode of Darwinian selective displacement would occur.
live longer than the actors of the competing S-lineage(s), then the
mortality (expected death rate) of the S-lineage will be less than
its competitors; or if the actors all uniformly have more
offspring (per unit time) then the fecundity (expected birth
rate) of the S-lineage will be greater than its competitors. In
either case, the size of the S-lineage would be expected to
increase relative to its competitor(s); and if the difference
were large enough, and maintained long enough, the S-lineage would
displace its competitors essentially deterministically - which is
to say, an episode of Darwinian selective displacement would occur.
This way of thinking leads to associating fitness with individual
actors, at least as a form of shorthand. This is always
legitimate in a certain abstruse, formal, sense. The members of
an S-lineage are guaranteed to be "similar" minimally in the
sense that they are all members of that S-lineage. So any
attribute of an S-lineage (such as fitness) can be imputed to
the putative "Similarity Class" (S-Class) which also
characterises all members of the S-lineage, and thus, implicitly,
to single actors, considered as "exemplars" of their
S-class.![]()
To put it in a slightly different way, any single actor is necessarily a member of its own S-lineage(s). Thus, it can surely favour its own S-lineage(s) by having as many offspring as it can (involving some kind of trade-off between living as long as it can while procreating as fast as it can). Again, this viewpoint estimates or measures fitness by reference to the activities or attributes of any single "typical" member of the S-class of actors making up a particular S-lineage.
The problem with this point of view is, however, that it is grossly simplistic. There may well be no such thing as a "typical" actor of a given S-lineage. In the simplest case, the selectively important "trait" may not be "expressed" at all in certain members of the S-lineage - but they can pass it on, and thus do indeed contribute to the selective dynamics of the S-lineage as a whole. But more generally, it is perfectly possible for the selectively important "trait" to be "expressed" in very different ways by different actors of the S-lineage. Thus, actors of the same S-lineage may be very different in a wide range of characteristics or attributes; or, for that matter, any single actor may engage in a wide variety of different behaviours, or manifest different traits, in different times and circumstances; we require only that these differences not be such as to establish two distinct, competing, lineages with distinct fitnesses relative to the particular selective competition we are studying (otherwise, of course, we would not be dealing with a single coherent S-lineage). The point is that the thing which is selectively important may not be, in any useful sense, a "trait" of a single actor at all, but rather is a "trait" of some groups of actors, or perhaps of the S-lineage as a whole.
Indeed, the fitness of an S-lineage may actually critically rely on the very distribution of variations in certain characteristics among the actors making it up. In that case, no single actor, qua actor, would even allow a determination of (S-lineage) fitness - except, perhaps, in the extremely contrived sense that we might, in theory, be able to somehow deduce or determine from that one actor what would be the "typical" distribution of variations within an S-lineage which it could found.
We can envisage a range of different ways in which variation in the characteristics of actors could be beneficial to an S-lineage. For example:
In general, both these possibilities could equally be based on dedicating certain actors to certain tasks on a lifetime basis, or on the use of a structured life cycle with different specialisations on the part of a single actor at different stages in its cycle; and, of course, the two possibilities could both be exploited to greater or lesser extents in a single S-lineage.
The notion of an actor being so constituted (in structure and/or behaviour) that it functions for the benefit of other actors, at a cost to itself (especially as measured in terms of the number of offspring it produces), is technically referred to as altruism, and has historically been viewed as somewhat problematic for Darwinian theory. However, we can now see that this derives from a blinkered view of selection: the view that supposes that an actor can benefit its S-lineage(s) (improve the latter's fitness) only through direct "benefit" to itself. In particular, the second scenario mentioned above, where an S-lineage can benefit from mutual co-operation between its members, clearly allows the possibility of altruism (on the part of actors to other actors in a single lineage) being positively favoured by a Darwinian selection process.
I have suggested that altruism could be favoured where there is a mechanism for actors to recognise other actors sharing a single S-lineage. It seems to me that this is a necessary condition. By definition, altruism involves costs to the actors engaging in it. This can conceivably provide a net benefit to the S-lineage only if most of the benefit of the altruism is retained within the S-lineage. To put it another way, altruism is in permanent danger of being subverted or exploited. If one S-lineage contains actors which are unconditionally altruistic, and another is otherwise similar (and thus competing) but its actors succeed in restricting their altruism (even with limited effectiveness) to members of their own S-lineage, then the latter S-lineage clearly gets more benefits from altruism than its competitor, and will be favoured by selection.
Note that this argument would not generally go though if we
considered a putative S-lineage which abandoned altruism
entirely. Such an S-lineage could initially grow
at the expense of the original S-lineage of altruistic actors.
But recall our basic hypothesis that the altruism of the actors
was indeed beneficial to the S-lineage. The new S-lineage, of entirely
selfish actors, would, as it becomes more numerous, start
losing this benefit. There is no general answer as to the final
outcome of such a process, but it certainly does not follow that
the original S-lineage would necessarily be
eliminated.![]() It would be equally possible that another different S-lineage
would arise which restricted its altruism: this would
then generally be capable of selectively eliminating both
of the others.
It would be equally possible that another different S-lineage
would arise which restricted its altruism: this would
then generally be capable of selectively eliminating both
of the others.
So altruism, if it arises, is expected to rely, to a greater or lesser extent, on mutual recognition between the actors of a single S-lineage. There are two basic mechanisms which suggest themselves for such recognition:
The former corresponds to what is conventionally referred to as "kin selection" in biological Darwinism, (Dawkins 1979). The latter is not generally explicitly emphasized in biology, though Dawkins has considered a variety of mechanisms which might come under this general heading (Dawkins 1982a, Chapter 8). Notwithstanding this apparent lack of emphasis among biologists, I speculate that benefits of altruism (in the broadest possible sense) mediated by recognition of class and/or recognition of close kin may be quite significant for Darwinian evolution generally. I say this simply on the basis that mutual recognition of this sort is clearly, in the general case, a difficult problem that calls for complex sensorimotor co-ordination, i.e. the utilisation of relatively complex anticipatory models of the world. Benefits (for S-lineages), deriving from more and more effective recognition and discrimination, seem to provide one plausible general basis for a sustained correlation between fitness and knowledge (or "adaptive complexity" ), and thus for an extended period of growth of knowledge through evolutionary time. However, intriguing as this possibility may be, it is of limited relevance to my purposes here, and I shall not pursue it further.
To return to the central idea of this section, which is the notion of the S-lineage as the unit of selection, the significance of this point can be put at its most stark by asking what is the "selective" value to an actor of procreation itself? Actually, there is none, and yet everything else that might be an adaptation of an actor is typically identified in terms of its contribution to procreation. Conversely, there is no problem when we think of the structure and behaviour of actors, insofar as it has been adapted by Darwinian evolution, as being consistently for the benefit of the S-lineage (procreation of at least some of the constituent actors is an absolutely necessary condition for the continued existence of any S-lineage). Which is simply to say that S-lineages which have come about through Darwinian evolution, are expected to be characterised by naked and exclusive selfishness; altruism on the part of an S-lineage (as opposed to the part of its component actors) could only be classified as pathological in a Darwinian context.
The essential point is that while the adaptations of an S-lineage may be clearly manifested in "adaptations" of the actors, this is an incidental effect, which may or may not occur, rather than being an intrinsic feature of Darwinian evolution in general. The general and reliable feature is the selfishness of the S-lineages themselves. The failure to realise this is manifested in the attempts to introduce entirely counter-intuitive modifications to the notion of "fitness" (of an actor) when it is discovered (as, for example, in the case of kin selected altruism) that, without such a correction, entities of lower "fitness" are apparently favoured by selection. This is the general idea of so-called "inclusive fitness" . Dawkins has rightly pointed out that that is an inspired, but ultimately misguided, stratagem (Dawkins 1978a): the supposed difficulty simply does not arise when one realises that the actors are not the appropriate entities to ground the analysis in the first place - the S-lineages are.
So I consider that the unit of selection is an S-lineage; and that the implication of this is that a given phenomenon can be considered as an outcome of Darwinian evolution if and only if it is consistent with an unconditional selfishness on the part of the S-lineage possessing or exhibiting it. This, in short, is the doctrine of The Selfish S-lineage.