
So: Does it or Doesn't it?
 The Elusive Unit of Selection
The Elusive Unit of Selection
The biological literature on the Units of Selection issue also addresses another question, quite different from the one I have just chosen to answer. This second question might be phrased as follows:
Is there a uniquely distinguished "level" of organisation (biological or otherwise) which characterises entities that can qualify as Darwinian actors?
I suggest that this is precisely the question which Hull set out to answer; whereas it is not clear to me whether Dawkins has ever consciously recognised this second question - not, at least, as a separate question. However he does seem to suggest an answer (implicitly or otherwise) - namely that genes are uniquely distinguished as the only biological entities which should properly be said to be actors. I believe that that claim is quite mistaken (but also that it is questionable whether Dawkins ever really intended to make it).
Specifically, to the extent that we interpret "gene" as a material part of an organism (a fragment of DNA, say), then I accept that such genes might be usefully considered as actors (in "suitable" circumstances); but I reject absolutely the idea that they are uniquely qualified for this role. I suggest that there are always alternative candidate actors (particularly, but not exclusively, organisms); alternatives which will be equivalent in the precise sense of yielding a selection dynamics which is either identical, or differs only by a bijective transformation of the state variables. Preferences among these different candidates therefore arise only from pragmatic considerations relating to the particular circumstances in which the theory is being applied.
It is, of course, no accident that "replicators" or "genes" turn up in two quite different senses in the two distinct questions regarding "units" of selection. For Dawkins, genes are the prototypical examples of replicators - and his conflation of both actor and S-lineage into the single term "replicator" is more or less mirrored in his usage of the term "gene" . Again, we should try to distinguish between (at least) actor-genes or A-genes, and lineage-genes or L-genes. The former may (or may not) play the role of actors, and, correspondingly, the latter may (or may not) play the role of S-lineages. Dawkins effectively argues, correctly in my view, that only S-lineages can qualify as units of selection. But he also equates this with a claim that only "genes" (or "replicators" ) can qualify as units of selection; I can accept this also, if it is read in the sense of "only L-genes (as opposed to A-genes)" or "only L-replicators (as opposed to A-replicators)" . But Dawkins seems to go on to parlay this into a claim that only "genes" - now specifically meaning A-genes, as opposed to (say) "organisms" - can qualify as Darwinian actors. I believe this to be definitely in error.
To repeat: I reject the idea that a certain level of biological organisation - the "genes" or the "DNA" or the "genetic material" or "replicators" - uniquely fulfills the role of Darwinian actor in terrestrial evolution. I refer to this flawed idea as genic selectionism - as opposed to organismic selectionism (Wright 1980, McMullin 1992b). My position is the pluralist one that genic and organismic selection, properly viewed, are not competitors or rivals, but merely alternative, formally interchangeable, descriptions of the same underlying biological reality; that, indeed, there may exist an indefinite number of other, similarly equivalent, descriptions; but that, whichever of these viewpoints may be adopted, there will be a crucial distinction between actors and S-lineages, with only the latter being properly regarded as units of selection, or entities for whose benefit (Darwinian) adaptations may be said to exist. This latter question, of actor versus S-lineage as the unit of selection, thus cuts at right angles to the question of gene versus organism as Darwinian actor; confounding these two questions leads only to confusion and error.
I may note that I do hold that there is something special, in evolutionary terms, about the "gene" level; or, more precisely, about that kind of actor organisation which involves a functional separation into a relatively passive information carrier or "genotype" , and a relatively active information processor or "phenotype" (Fig.). This is, of course, the von Neumann architecture for self-reproduction (Von Neumann 1951, 1966a, 1966b), and it exhibits features which are crucially important for the possibility of a sustained Darwinian growth of complexity. Strangely, that this is so, and that this is precisely why von Neumann proposed the architecture he did, is still not generally appreciated even now, almost half a century after his seminal investigations.
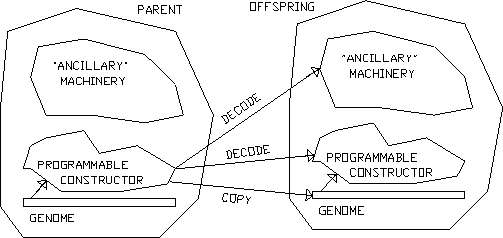
Figure 3: "Genetic" actor organisation.
Unfortunately, a properly detailed analysis and critique either of Dawkins' (alleged) genic selectionism, or of von Neumann's general theory of automaton evolution, would take me much too far afield in the current context. The interested reader (!) can find these discussions fully elaborated in (McMullin 1992c) and (McMullin 1992d, Chapter 4), respectively.
So: Does it or Doesn't it?
The Elusive Unit of Selection