Preliminary Steps toward
Artificial Protocell Computation
- DCU Alife Lab:
- Ciarán Kelly
- Darragh O'Brien
- George G. Mitchell
- James Decraene
- Financial support under EU FP6 Integrated Project PACE
Contract number: 002035
- Metabolism
- Containment (membrane)
- Information (heredity?)
- It might be at the (proto)-cell level:
- Cell cycle control
- Chemotaxis
- (Differentiation? Apoptosis?)
- ...
- It might be at the multi-(proto)-cell level:
- Nervous system
- Immune system
- ...
We will concentrate on the single (proto-)cell level.
- Examples?
- Cell signalling networks, CSN
- Genetic regulatory networks, GRN (beyond protocells?)
- Molecular Information Processing:
- Operators: catalysts/enzymes
- Operands: substrates, reactants
- Real time
- Reaction network -- somewhat like term re-writing system
(but no demarcation between rules and messages)
- Reaction network ``closure'' matters (why?)
- Protocell as encapsulated ``replicator world''?
- Concentration matters (as does stoichiometry, thermodynamics,
kinetics, catalysis ...)
- Evolution
- At the protocell level
- Layered on ``replicator'' dynamics at the molecular level
- (AKA ``major transition'')
- Work with an ``artificial chemistry''
- Polymer family composed of two categories of monomer:
labelled 0 and 1 (primary structure is binary
string)
- No thermodynamics (!)
- No material conservation (!!)
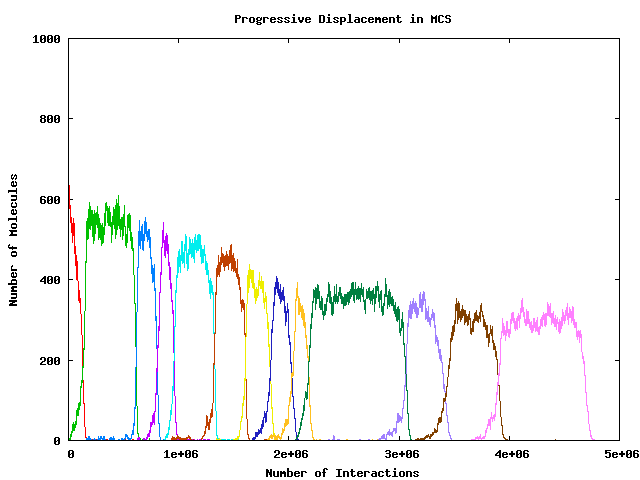
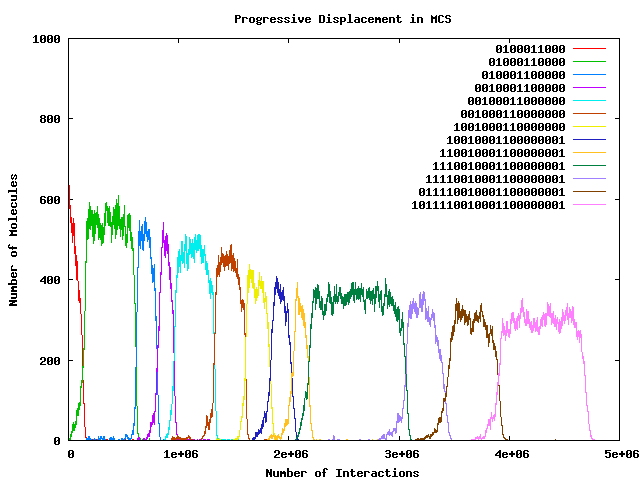
- Why does dominant string not take over whole population?
- What is the composition of the rest of the population?
- Why does peak dominant population get progressively smaller?
- (... obviously there is some stuff that has not been
explained yet!)
- In this particular ``flow reactor'' system,
longevity and fecundity are exactly, inversely,
coupled, so no evolutionary scope there.
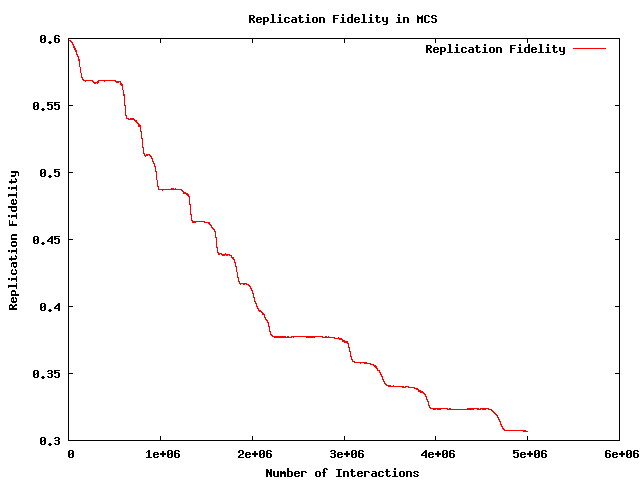
- So: that leaves fidelity -- doesn't it?
- (Think of this as a simple ``experimental control'' or
``sanity check''.)
- Only difference in ``intrinsic'' fitness of the different
replicator species is in ``fidelity'' - and this shows a
progressive, quasi-deterministic, decay.
- Along with this goes reducing concentration of dominant species
(increased mutational load) and consequently reducing fecundity
(albeit with exactly inversely increasing longevity).
- Long term outcome is total distintegration of the original
organisation (progressive - no ``threshold'' effect).
- But this is happens through a sequence of ``short term''
events, each showing perfectly ``darwinian'' selection!?
- Enzymatic ``binding rule'' is ``exact substring match'' (Aaahhhh ...)
- So a ``super-string'' will immediately parasitise any host
sub-string which was previously dominant (think ``hyperline'' rather
than ``hypercycle''!); and will quickly displace it completely.
- All the ``selectional'' events in this particular model are
of this nature: the huge ``parasitic'' gain easily outweighs
each slight, incremental, decrement in fidelity (reducing
intrinsic fitness).
- Playing games, not climbing mountains (improbable or otherwise!).
- Bimolecular, ``self-catalysed'', replication has hyperbolic
growth rate.
- Implies density-dependent selection with positive
feedback.
- Result is ``survival of the common'': invasion is extremely
difficult - even by rivals with much higher intrinsic fitness.
- Very different from classical ``auto-replication''
(mediated by an externally buffered replicase) with exponential
growth rate and yielding straightforward Darwinian selection.
- But that's a different story ...
- ``Yes, it's a little counter-intuitive ...
- ... but it's really just a very very contrived
and peculiar toy system, with no wider ramifications!''
I may not have gone
where I intended to go, but I think I have ended up where I
needed to be. -- Dirk Gently
- At the very least, it underlines that the Dawkins' slogan
(``Longevity, Fecundity, Fidelity'') is wildly over-simplistic.
- We suggest that it was worthwhile to isolate and
characterise this phenomenon clearly before adding
additional complications.
- But best of all: it immediately offers a simple
candidate problem for solution by protocell level selection.
- So stay tuned!
- Presentation slides:
- DCU Alife Laboratory:
- Research Institute for Networks and
Communications Engineering (RINCE):
Copyright
This work is copyright ©2007 by
Barry McMullin.
Permission is hereby granted to private individuals to access,
copy and distribute this work, for purposes of private study
only, provided that the distribution is complete and unmodified,
is accompanied by this copyright notice, and that no charges are
levied. The work may not be accessed or copied, in
whole or in part, for commercial purposes, except with the prior
written permission of the author.
All other rights reserved.
Copyright © 2007 All Rights Reserved.
Timestamp: 2007-03-26
Barry.McMullin@dcu.ie